ESTUDIO SOBRE EL SONAR DE LOS MURCIÉLAGOS
Y SU UTILIZACIÓN EN LA CAZA
En esta sección se han recogido los aspectos más interesantes de un estudio realizado en Maryland sobre el haz emitido por el sistema sonar de los murciélagos a la hora de cazar y rastrear a sus presas (ver bibliografía). La fuente de este informe es:
"The sonar beam pattern of a flying bat as it tracks tethered insects"
Kaushik Ghose and Cynthia F. Moss
Neuroscience and Cognitive Science Program, Department of Psychology, The University of Maryland,
College Park, Maryland 20742
~Received 31 July 2002; revised 25 April 2003; accepted 12 May 2003
El informe describe la forma y características del haz del sonar de los Eptesicus fuscus, murciélagos que utilizan la ecolocalización para orientarse. Para ello se realizan varias capturas de insectos en un gran laboratorio que sirve como "habitación de vuelo".
El "modelo de haz" (beam pattern) se deduce a partir de la intensidad de la señal emitida, la cual se mide utilizando un array linear de micrófonos. Las posiciones del murciélago y su presa se obtienen a través de una reconstrucción estereoscópica de las vistas de dos cámaras.
Los resultados se muestran de forma gráfica y estimando la dirección del eje del rayo (beam axis). El murciélago centra el eje del haz sobre el objetivo seleccionado con una desviación estándar de 3º. El error experimental es 61.4º.
Los ensayos dirigidos con dos objetivos muestran que el murciélago se centra en uno de los blancos y lo rastrea con su sonar. Estos hallazgos muestran que el eje del "haz de señales acústicas" que emite el murciélago es un buen indicador de rastreo selectivo de presas, análogamente a la forma en que persiguen su objetivo los animales "visuales" (fijando la mirada en la presa).
I. INTRODUCCIÓN
Los murciélagos pueden orientarse, buscar comida y desarrollar otras tareas perceptivas guiadas en completa oscuridad mediante la emisión de señales de voz ultrasónicas y analizando los ecos que vuelven procedentes de los objetos de su entorno. En este sentido, los murciélagos nos dan la oportunidad de estudiar el uso de la audición en tareas espaciales, en las que otros animales utilizan la visión.
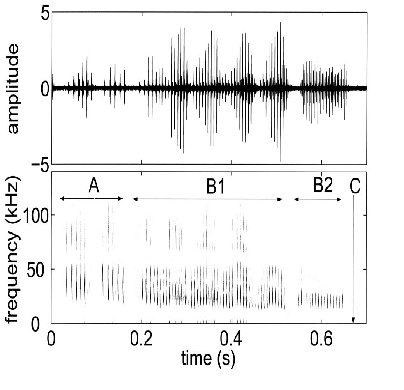
Estudiaremos el Eptesicus fuscus, una especie de murciélago que se ecolocaliza con señales de frecuencia modulada (FM). Cada señal de sonar consiste en varios barridos de frecuencias relacionadas armónicamente. El tiempo de duración de la llamada en la ecolocalizacion para el E.fuscus – la estructura en frecuencia - cambia según si el murciélago está buscando, acercándose o capturando a su presa (fig.1). Esta especie busca comida principalmente en espacios abiertos, aunque se ha visto que también persiguen a sus presas cerca de vegetación.
El ritmo, duración y características espectrales de cada pulso del sonar influye en la información del sistema de imágenes acústicas del que dispone el murciélago. Mientras busca a su presa, el murciélago utiliza pulsos largos (15 a 20ms) con barridos de frecuencia superficiales. La frecuencia fundamental abarca de 28 a 22KHz aproximadamente. La tasa puede ser aproximadamente de frecuencias tan bajas como 5-10Hz. Una vez que detecta una nueva presa, el murciélago se acerca a ella, acortando los pulsos a 2-5ms y aumentando el ancho de banda (normalmente barriendo de 60 a 22KHz). Durante la fase terminal los pulsos pueden llegar a ser de 0.5-1ms, con el barrido fundamental de 40 a 12KHz y producido a tasas de hasta 150-200Hz en el último zumbido (ver fig1). Las vocalizaciones cesan cuando el murciélago se encuentra a 10-15cm de su presa (que es aproximadamente 30-50ms antes de poder estar en contacto con ella).
La secuencia se completa con el intento de captura utilizando la cola-membrana (colocándola como una cuchara), las alas inclinadas (para empujar la presa hacia la boca), o, en casos excepcionales, atrapándola directamente con la boca. Las señales de búsqueda de mayor duración se han grabado solamente en espacios abiertos, no en laboratorios.
Las características espaciales del haz del sonar también influyen en los ecos recibidos por el murciélago. Se ha analizado que el haz del sonar es ancho, pero no omnidireccional. Asimismo, se ha experimentado que el murciélago dirige sus vocalizaciones hacia la presa para intentar maximizar la relación señal a ruido de los ecos (los objetos próximos al eje del haz devolverán ecos de mayor intensidad que los que se encuentran más alejados).
FIG. 1. La imagen superior muestra la forma de onda en el dominio del tiempo de una serie de vocalizaciones del E.f.uscus grabadas en un laboratorio. La imagen inferior muestra el espectograma de esta señal. En ella se señalan las diferentes etapas de la búsqueda de alimento. A es la fase de aproximación, B1 es el primer zumbido, B2 el Segundo y C se refiere al momento en que el murciélago contacta con su presa.
Por lo tanto, se deduce que el objetivo de orientar el sonar del murciélago en la dirección de la presa es una acción motora natural asociada con la selección del objetivo y el rastreo. Esta hipótesis se demostró grabando el haz del sonar (sonar beam) producido por los murciélagos cuando capturaban insectos atrapados en el cuarto de vuelo. Se usaron estos datos para calcular la dirección del eje del haz (beam axis) en relación con el objetivo.
II. MÉTODOS
A. Ensayos en el laboratorio.
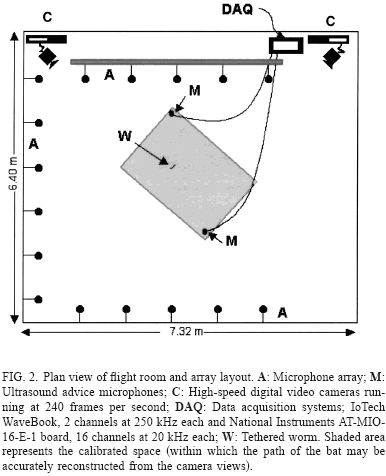
Se utilizaron cuatro murciélagos E.fuscus para el estudio. Los murciélagos se soltaron en un cuarto grande ~736 m2, cuyas paredes estaban recubiertas de espuma absorbente de sonido para evitar la reverberación y permitir las grabaciones de las vocalizaciones del murciélago. Los murciélagos se entrenaron para atrapar un gusano enganchado en un soporte mientras volaban. El objetivo podía moverse en un camino circular gracias a un motor situado directamente en el techo. También podría dejarse caer en el espacio de vuelo por un mecanismo de trampa-puerta instalado en el techo. La puerta de la trampa se forró para minimizar el ruido cuando se abriese. Los micrófonos puestos en el suelo del cuarto no recogían ningún sonido cuando la puerta de la trampa se abría.
Uno de los experimentos sobre el comportamiento consistía en soltar el blanco atado desde la puerta de la trampa en un punto aleatorio cuando el murciélago pasa volando por él. De esta forma el murciélago se encontraba con un objetivo cuya localización (en un área de aprox. 4m2) era totalmente desconocida hasta que se abría la puerta de la trampa. El análisis de la dirección de las emisiones antes y después de que apareciese la presa permitió estudiar varios aspectos de la conducta del murciélago cuando detecta y ataca a sus presas. Los cuatro murciélagos habían sido entrenados previamente para atrapar blancos de un soporte y se habían usado el año anterior para estudiar sus vocalizaciones al coger insectos atados en el laboratorio. Los resultados se recogían en cada experimento de captura del insecto, en forma de segmentos de datos que contenían uno o más intentos del murciélago de capturar el objetivo.
B. Arrays de grabaciones.
El array estaba formado por 16 micrófonos colocados en forma de U a lo largo de las tres paredes de la habitación de vuelo se alojan ~ver Fig. 2. El espacio lineal entre los micrófonos era 1 m, y la altura de los micrófonos era 0.9 m sobre el suelo. Estaban situados de forma que se redujese el solapamiento entre el sonido original y cualquier eco residual.
Son numerosos los experimentos que se han realizado con murciélagos en laboratorios que disponen de paredes insonorizadas, para evitar producir cualquier tipo de sonido excepto el de una presa "ficticia". La información se recoge en forma de segmentos de datos que reflejan uno o más intentos del murciélago para capturar el objetivo. Para poder estudiar el "beam pattern" para una banda de frecuencia dada necesitamos conocer la intensidad de la señal en esa banda. Esta información puede obtenerse tanto de la señal paso banda como de la envolvente de dicha señal, asumiendo que la señal es de banda estrecha (o si no descomponiéndola en segmentos en los que lo sea).
Si f(t) es la señal medida, fa(t) será la señal analítica de f(t) y f^(t) su transformada de Hilbert. Así:
Sabemos que la envolvente de f(t):
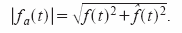
Por lo tanto, la integral del cuadrado de la envolvente entre los instantes t1 y t2 se reduce a:
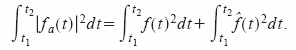
El primer término
de la expresión anterior corresponde a la energía de la señal en el periodo de
tiempo comprendido entre t1 y t2. Si asumimos que la señal en este período de
tiempo tiene una sola componente frecuencial, f^(t) sera simplemente una versión
de f(t) en la que lo que varía es la fase. Si además asumimos que el intervalo
t1-t2 es mucho mayor que el periodo de la señal f(t) en el que se cumple que
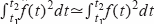 ,
obtenemos:
,
obtenemos:
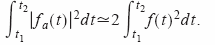
Este resultado muestra que integrar el cuadrado de la envolvente de la señal paso banda originada por el murciélago nos permite obtener la intensidad de la señal en esa banda. Las simulaciones utilizadas de vocalizaciones de murciélago grabadas confirman este resultado. Como ya se ha mencionado. Las vocalizaciones de sonar del E.fuscus son barridos de frecuencia compuestos por una frecuencia fundamental y varios armónicos.
El contenido en frecuencia de la envolvente para las señales de ecolocalización está relacionado con la duración de las señales. La parte de menor duración de la señal corresponde con la última fase de la captura del insecto, en la que los "zumbidos” son del orden de 0.5-1ms, lo que indica que el límite superior de frecuencia de toda la envolvente de la señal será aproximadamente 2KHz. Si suponemos además que la señal paso banda tiene una duración de 0.25ms, los 2KHz se "trasladaran" en frecuencia hasta 4KHz aproximadamente. Por lo tanto, una tasa de muestreo de 20KHz capturará la envolvente con buena fidelidad.
El contenido en frecuencia de las señales de sonar del E fuscus cazando insectos en el laboratorio varía mucho, dándose frecuencias más altas durante el primer acercamiento a la presa y frecuencias más bajas durante los últimos zumbidos. Eligiendo una frecuencia central de 35KHz descubrimos que se pueden grabar señales de todas las fases de al búsqueda de alimento. La SNR típica de las vocalizaciones del murciélago para este método era de 20dB.
Para determinar el "beam pattern" de un murciélago volador, la atenuación en función de la distancia de las señales debe ser corregida para realizar buenas mediciones. Esta corrección tiene dos componentes. Uno es la atenuación esférica y depende sólo de la distancia entre el murciélago y el micrófono. La otra es la absorción de energía conforme el sonido se propaga por el aire. Esta depende tanto de la frecuencia como de la distancia.
Para solucionar esto se desarrolla el esquema siguiente:
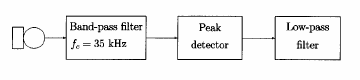
La señal procedente de cada micrófono pasa por un filtro paso banda que la amplifica y extrae los componentes entorno a la frecuencia central (35KHz). Las características en frecuencia de este filtro son:
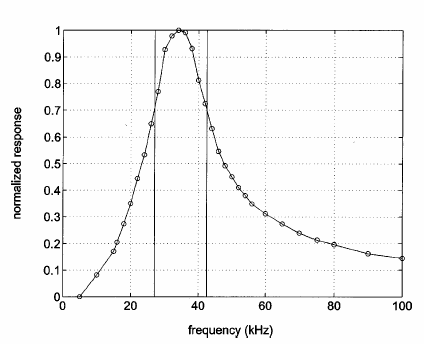
FIG. 3. ~A! Schematic of signal-processing hardware.! Filter characteristics of the bandpass filter used. The x axis shows the frequency, while the y axis shows the normalized response. The vertical bars correspond tothe 3-dB ~half-power! points, i.e., the start and stop frequency.
Posteriormente, la señal pasa por un circuito detector de pico que extrae la envolvente de esta señal paso banda. Esta envolvente se suaviza con un filtro paso bajo y después se digitaliza. Veamos algunos ejemplos de señales de sonar de murciélagos recibidas en el micrófono cuya envolvente es filtrada paso banda y suavizada:
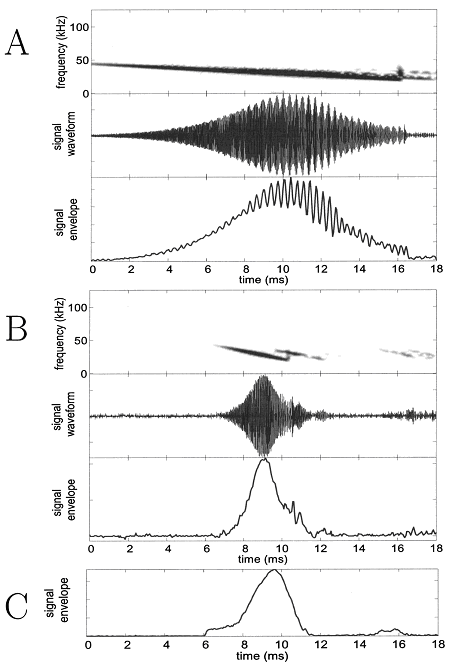
FIG. 4. Panels ~A! and ~B! show recordings taken by pinging the array with frequency sweeps from an emitter. A shows that for a long ~shallow! sweep there is more overlap, between the incident sound and the returning echo, and the beats are more prominent. B shows that for short sweeps there is less overlap. The top panel in each is the spectrogram of the bandpassed signal received at one of the array microphones, the middle panel shows the time waveform of that signal, while the bottom panel shows the envelope extracted by the array hardware. The interaction between the incident sound and an overlapping echo shows up as a beat. In both A and B, the emitter was placed in the plane of the array so as to maximize the echo returning to the microphone from the array backend. Due to limitations of the signal generator used to produce the emitted sounds, each frequency sweep has a brief glitch as it resets to the start frequency and this is visible as a vertical streak in the spectrogram. This does not change
any results. ~C! shows the envelope signal taken from an array circuit during a trial with a flying bat. In general, the bat sounds recorded at the array do not show apparent effects of overlapping echoes.
III ANÁLISIS DE LOS DATOS.
A. Cálculo del "beam pattern".
Como ya se ha señalado, las señales recibidas se segmentan para seleccionar las vocalizaciones y excluir los ecos. La intensidad recibida, Ir, se calcula a partir de la envolvente. Se corrigen este valor teniendo en cuenta las pérdidas esféricas y la atenuación atmosférica, obteniendo finalmente Ic. La intensidad corregida Ic se calcula a partir de la fórmula:
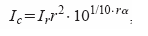
,donde "r" es la distancia entre el micrófono y el murciélago.
El "beam pattern" global se muestra en la fig.5:
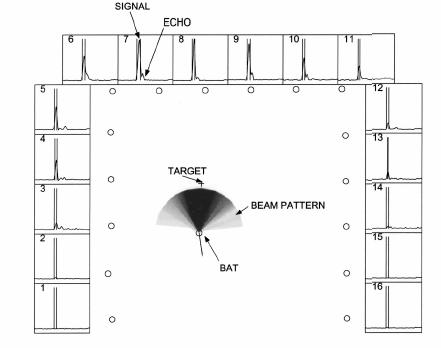
FIG. 5. Beam-pattern reconstruction. Central panel shows the reconstructed beam pattern. The 16 circles along the edges of the panel are the positions of the microphones. The pattern is normalized such that the peak intensity has a value of 1.0 and is colored black. Lighter colors denote progressively lower intensities. The circle at the center of the beam pattern represents the position of the bat. The 1 symbol represents the position of the worm. The thin curved line terminating at the bat’s position is the trajectory of the bat up to that frame. The straight line drawn from the bat represents the direction of motion of the bat ~in this frame the two overlap!. Surrounding panels ~numbered 1 through 16! show the envelope signals digitized from each microphone. All the side panels have the same scales. Twenty ms of data are shown. The signal on each panel is time shifted to compensate for the time of travel of the sound from the bat to the corresponding microphone. As a result the direct signal from the bat ~first sound! lines up on all the panels. A fairly loud echo ~second smaller bump! does not as its source is at a different position. This makes it easier to segment the signals and discard the echoes. The segmentation for the vocalization shown is depicted as two vertical bars bracketing the relevant portion of the envelope trace.
B: Cálculo del "beam axis"
Según las hipótesis anteriores, el murciélago dirige su sonar hacia el objetivo de interés. Asumiendo que el haz es simétrico, añadiendo vectores de dirección del murciélago a cada micrófono ponderados por la intensidad correcta, se obtiene un vector dirección que es una estimación objetiva del eje del haz, sin tener en cuenta el perfil real de éste.
Se ve en la siguiente ecuación:
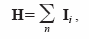
, donde Ii es el vector dibujado del murciélago al micrófono "i" con magnitud proporcional a la intensidad correcta. H es la resultante , cuya dirección es la estimación del eje del haz.
C. Errores debidos a la geometría del array de micrófonos.
La fig.6 muestra los resultados del cálculo del eje del haz para seis orientaciones de cabeza diferentes. El "beam pattern" simulado se muestra en el centro del array. Este haz se emite después en diferentes posiciones en el espacio, y las direcciones estimadas se calcularon de las señales recibidas de cada micrófono del array. Los resultados se muestran con flechas negras.
IV. RESULTADOS
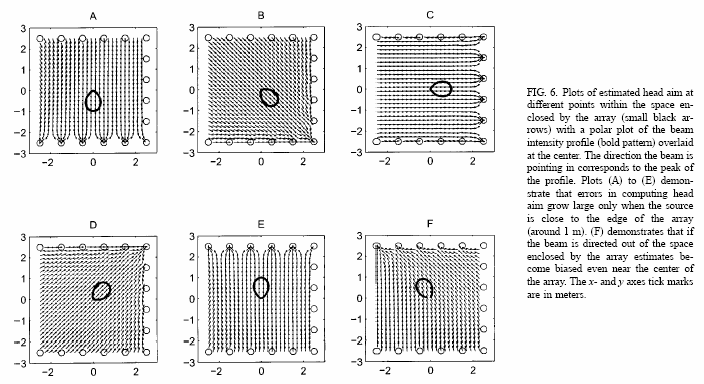
A. Beam patterns
El resultado principal del estudio son los "beam patterns", que se obtienen cuando el murciélago selecciona, rastrea y captura su objetivo. La fig.8 muestra una secuencia perteneciente a distintas vocalizaciones. Muestran claramente cómo el murciélago primero escanea el espacio que le rodea con su sonar (Figs. 8A, 8B y 8C) y posteriormente alinea su rayo con el objetivo. (figs. 8D, 8E y 8F). Hay que fijarse también en la muesca en los "beam patterns" A, B y D.
En el siguiente enlace se pueden encontrar animaciones y grabaciones de los experimentos descritos:
http://www.bsos.umd.edu/psyc/batlab/jasa03/
B.Anchura del haz
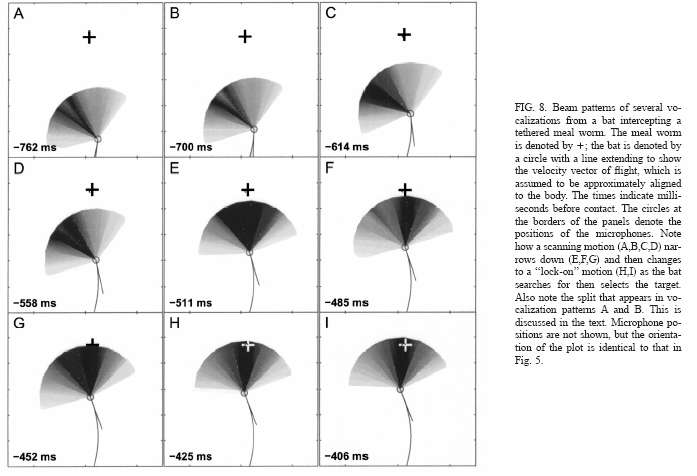
Las medidas fueron usadas para encontrar los puntos de potencia media del haz (aquellos en los que la intensidad es 3dB menor que la de pico).
la Fig.9 es un histograma en frecuencia de los 23dB de anchura del haz obtenidos por este método.
Los cuatro trazos corresponden a los datos de los cuatro murciélagos. La Fig.9B muestra un diagrama de dispersión de las anchuras del haz frente a la distancia del objetivo cuando fueron obtenidos. No hay ninguna correlación significativa entre la anchura del haz y distancia al objetivo (r520.0252, p.0.1). La mayoría de los puntos de datos se obtienen con el murciélago a menos de 1 m del blanco. El valor medio de 23dB para todos los murciélagos es 70º.
V. CONCLUSIONES
Estos experimentos son las primeras medidas del haz del sonar de los murciélagos cuando rastrean e interceptan presas en vuelo. Los datos presentados aquí sugieren que los murciélagos de la especie E. fuscus dirigen su haz al blanco de interés con una exactitud de aproximadamente 3º. Puede haber cierta analogía entre la orientación "acústica" del haz del sonar en los murciélagos y la orientación con la mirada de los animales "visuales" como los primates. Recientes experimentos sobre humanos han revelado que, ante la observación de una misma escena, el patrón de los movimientos del ojo está influenciado por la información que el sujeto está intentando obtener de la visión. La orientación de los ojos hacia los estímulos visuales es una acción natural muy importante, aunque los primates pueden, si es necesario, atender a cierto estímulo "secretamente", es decir, sin reposicionar los ojos.
Se ha visto que la orientación del haz puede usarse como un índice que revela algunos aspectos del estado interior del murciélago durante los diferentes ensayos de comportamiento. Específicamente, la orientación del haz puede usarse para comprobar qué objetos interesan al murciélago dentro de un entorno "complejo". También se ve que la conducta que sigue el murciélago al orientarse puede servir para medir latencias en las tareas de detección de objetivos por medio de la ecolocalización, muy similar a la utilización de los movimientos oculares en paradigmas visuales.